精神分裂症(Schizophrenia,SCZ)是一种高度复杂的神经精神疾病,其发病具有显著的遗传基础。佳学基因通过综合分析三类核心遗传学图形,系统阐述精神分裂症的遗传学机制。图一以等位基因频率与比值比(Odds Ratio)为坐标轴,揭示了不同类型基因变异对精神分裂症发病风险的贡献:罕见拷贝数变异(CNVs)如22q11.2缺失(OR>60)及罕见蛋白截短变异(PTVs)虽等位基因频率极低,却具有最高的个体致病效应;而单核苷酸多态性(SNPs)因频率高但效应量小,通过多基因累积方式贡献疾病风险。图二以嵌套环形图呈现精神分裂症遗传度的构成:疾病表型变异中80%具有可遗传性,其中SNPs贡献约25%的表型变异,CNVs与罕见编码变异(RCVs)各贡献约2%,而仍有51%的遗传变异尚未归因于已知变异类型,提示存在大量待发现的遗传因素。图三则从空间维度展示精神分裂症风险基因的突触定位:风险基因高度富集于突触后致密区(PSD)及突触后膜,关键基因包括GRIA3、GRIN2A、NRXN1、CACNA1C、DLGAP2等,涵盖谷氨酸受体亚基、突触支架蛋白及电压门控离子通道,提示谷氨酸能突触信号传导异常在精神分裂症发病中居于核心地位。综上,精神分裂症的遗传学病因呈现"共同变异-罕见变异"双轨模式,遗传效应汇聚于突触功能调控网络。佳学基因未来的研究会聚焦于未归因遗传变异的挖掘及突触蛋白互作网络的功能解析,以期为精神分裂症的精准诊疗提供理论依据。
关键词:精神分裂症;拷贝数变异;单核苷酸多态性;多基因风险评分;突触后致密区;谷氨酸能信号传导;遗传度;罕见变异
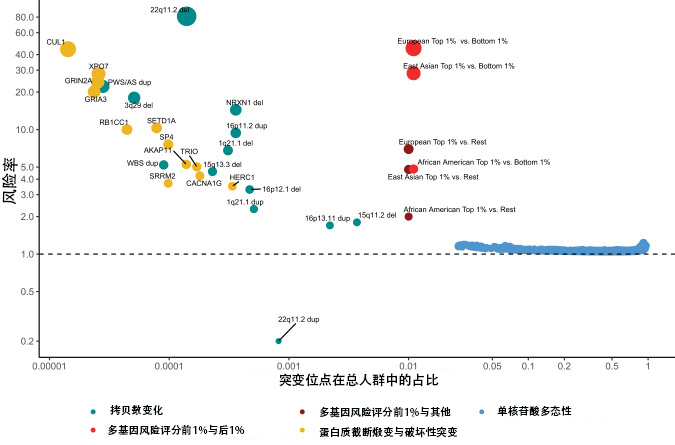
一、图形解析图一:不同基因变异对精神分裂症风险的效应比较本图以横轴表示一般人群中的等位基因频率,纵轴表示比值比(OR),通过散点大小与颜色区分五类遗传变异,揭示"效应量与频率的负相关规律"。
(1)罕见拷贝数变异(CNVs,青绿色)
代表性位点22q11.2缺失(DiGeorge综合征相关区域)的OR值超过60,是目前已知效应最强的精神分裂症单一遗传风险因素,但其等位基因频率仅约0.0001。其他显著CNVs包括:
NRXN1 缺失(OR≈10)
16p11.2缺失(OR≈8)
1q21.1缺失(OR≈6)
CUL1、GRIN2A、GRIA3相关区域缺失(OR 20–40)
值得注意的是,22q11.2重复(duplication)的OR约为0.2,提示该区域重复反而可能具有一定保护效应,与缺失方向相反,体现基因剂量效应的方向特异性。
(2)蛋白截短变异及有害错义变异(PTVs/damaging missense,黄色)
包括RB1CC1、SETD1A、SP4、AKAP11、TRIO、SRRM2、CACNA1G、HERC1等基因的罕见有害变异,OR值集中于3–10,频率约0.00001–0.001,属于罕见高效变异范畴。
(3)多基因风险评分对比(PRS,红色与橙色)
欧洲人群PRS最高1% vs. 最低1%:OR约为40–50,是迄今多基因风险最强的效应量证据
东亚人群PRS最高1% vs. 最低1%:OR约为30
非裔美国人群PRS最高1% vs. 最低1%:OR约为3–4,反映跨种族PRS迁移性的局限
PRS vs. Rest的比较(右上散点群)OR约为5–6,频率接近0.01,提示多基因风险在人群水平的显著累积效应。
(4)单核苷酸多态性(SNPs,蓝色)
分布于等位基因频率0.1–1.0区间,OR值极小(约1.0–1.1),紧贴虚线(OR=1基准线),体现"多个常见变异,每个效应微小"的多基因模型(polygenic model)核心特征。
核心规律: 遗传变异频率越低,个体效应越强;但高频SNPs通过数量累积同样可以产生重要群体风险。这一频率-效应权衡关系是精神分裂症遗传架构的基本特征。
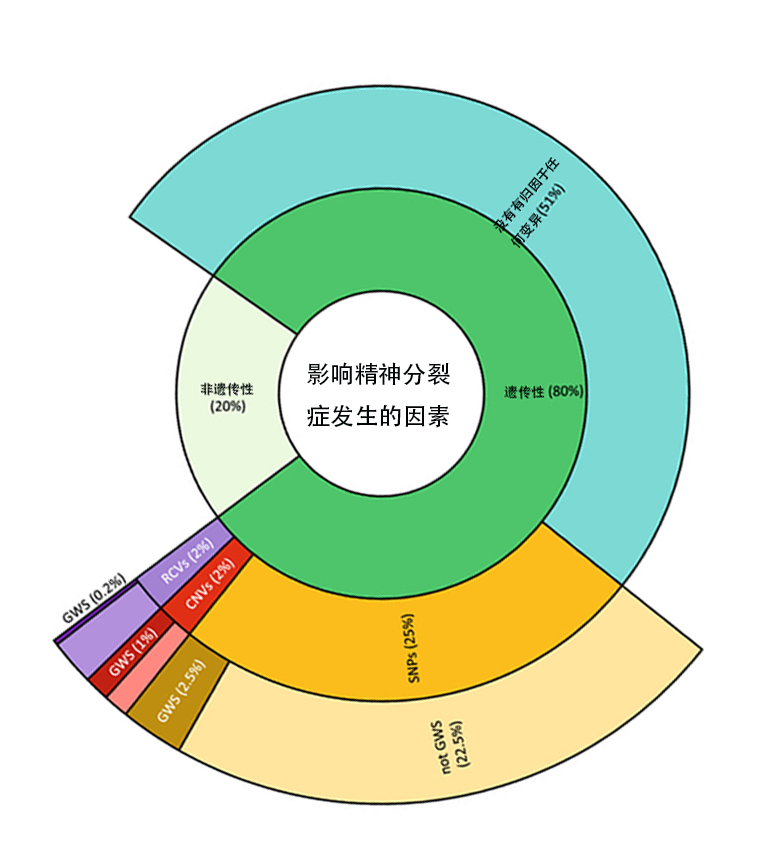
本图采用嵌套弧形图(sunburst diagram)从内到外分层展示精神分裂症表型变异的遗传学归因。
最内层——可遗传性划分:
可遗传部分(Heritable):80% 的精神分裂症表型变异具有遗传学基础,与双生子研究估计的遗传度高度吻合
非遗传部分(Not-heritable):20%,反映环境因素(围产期感染、城市化、大麻暴露等)的贡献
中间层——归因与未归因:
未归因于已知变异类型(Unattributed to variant type):51% 这是"遗传力缺失"(missing heritability)问题的直观体现,提示现有全基因组关联研究(GWAS)和测序技术尚未捕获的遗传信息(如表观遗传变异、基因-基因互作、结构变异等)
SNPs贡献:25%,为最大的已归因成分
CNVs:2%
RCVs(罕见编码变异):2%
外层——全基因组显著性(GWS)细分:
SNPs中达到全基因组显著水平(GWS)的变异贡献约2.5%,未达显著阈值(not GWS)的SNPs贡献22.5%,说明绝大多数SNP遗传信号尚处于亚显著水平
CNVs中GWS部分约1%,RCVs中GWS部分约0.2%
启示: 精神分裂症存在严重的"遗传力缺失"问题。扩大样本量、整合多组学数据及发展新型统计方法是揭示未归因遗传变异的关键路径。
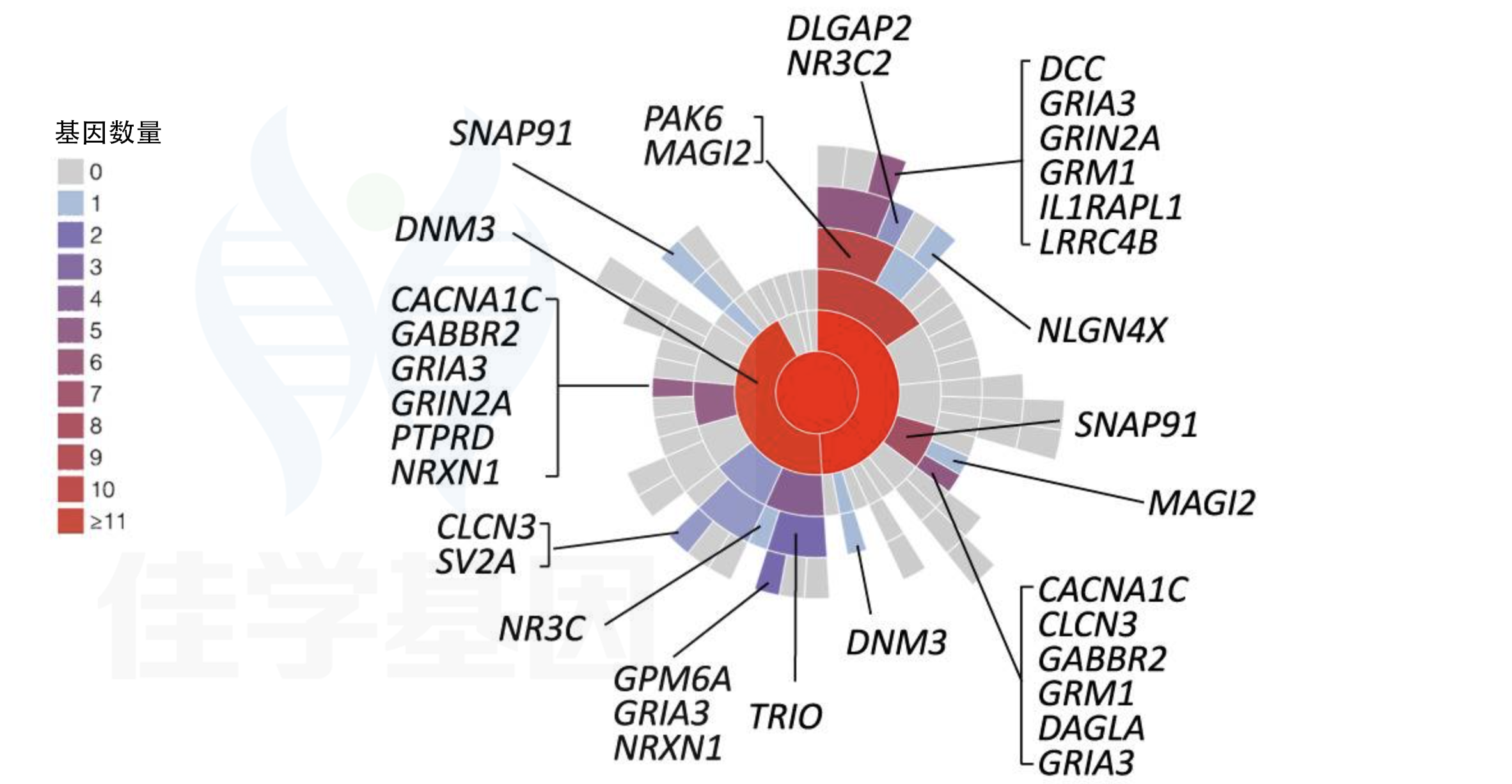
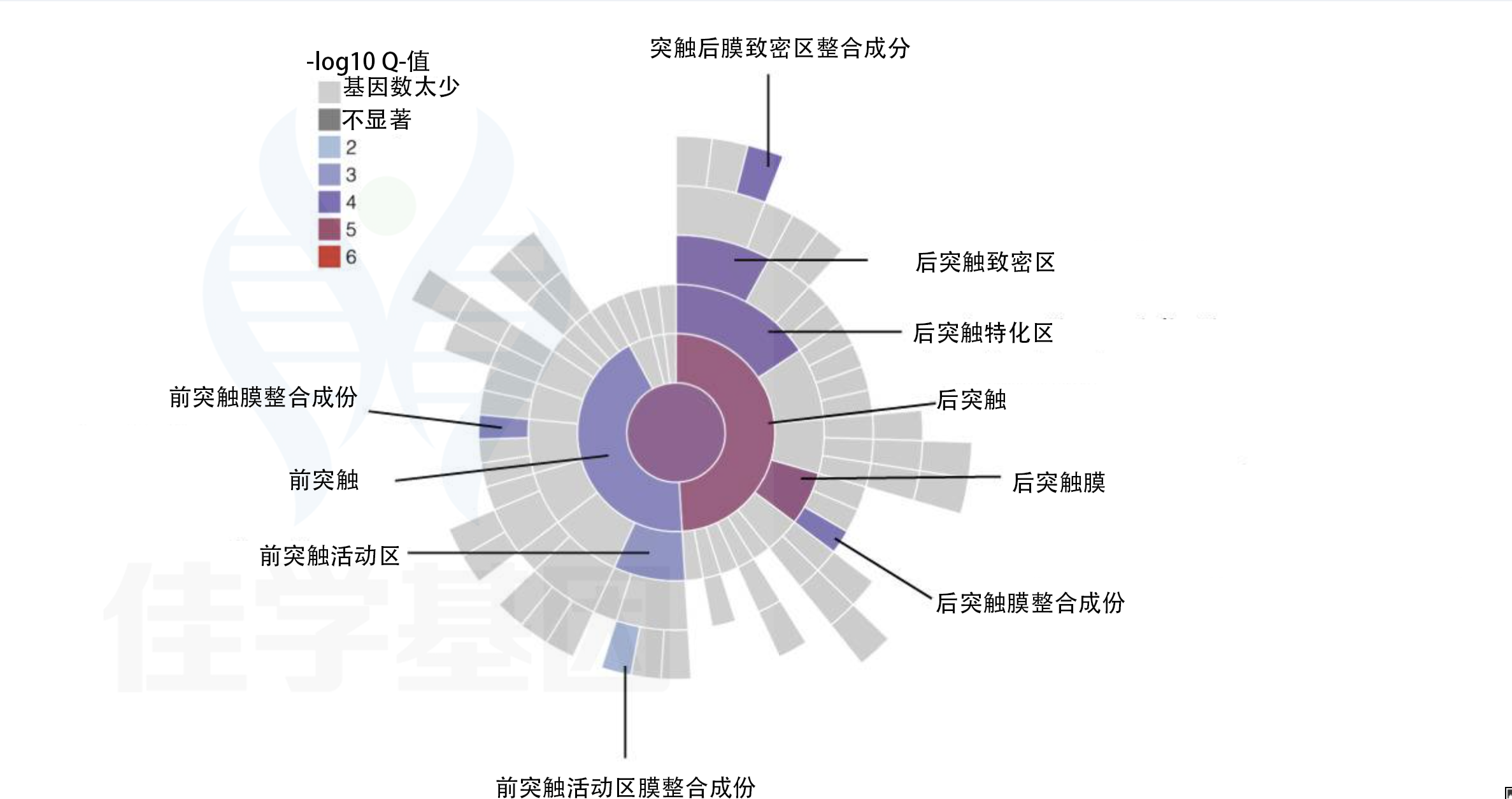
本图分为两个部分,采用放射状层级图(radial hierarchy plot)展示风险基因在突触结构中的空间分布规律。
A部分——突触定位的基因密度(gene count):
颜色越深(红色)代表该突触区室中富集的精神分裂症风险基因越多。图中清晰显示:
突触后致密区(PSD)及突触后膜是风险基因最密集的区域,代表性基因包括:
DLGAP2、MAGI2(突触支架蛋白)
GRIA3、GRIN2A、GRM1(谷氨酸受体亚基:AMPA/NMDA/代谢型)
NRXN1(突触黏附分子)
IL1RAPL1、LRRC4B(突触发育相关)
NLGN4X(神经连接素,与突触形成密切相关)
突触前膜亦有显著富集:
SNAP91(网格蛋白介导的囊泡内吞)
DNM3(动力蛋白,调控突触囊泡循环)
SV2A(突触囊泡糖蛋白)
CACNA1C(L型电压门控钙通道,调控神经递质释放)
B部分——突触层级组件的富集显著性(-log10 Q值):
颜色越深代表Q值越小(富集越显著)。结果显示:
**突触后致密区(postsynaptic density)和突触后特化区(postsynaptic specialization)**的富集显著性最高(Q值<10⁻⁶)
**突触后膜(postsynaptic membrane)和突触后完整膜组分(integral component of postsynaptic membrane)**同样高度显著
突触前活性区(presynaptic active zone)也有中等程度富集,但显著性低于突触后区室
核心结论: 精神分裂症风险基因在突触后谷氨酸能信号传导复合体中高度聚集,支持NMDA受体功能低下(NMDA receptor hypofunction)假说,并将突触可塑性损伤确立为精神分裂症的核心病理机制之一。
精神分裂症的遗传度约为60%–80%(双生子研究),属于高度多基因遗传疾病。其遗传架构符合"共同疾病-罕见变异"与"共同疾病-共同变异"的双重模型,即疾病风险由数量众多的常见低效SNPs与数量较少的罕见高效变异(CNVs、PTVs、错义突变)共同决定。
2.2 常见变异:多基因风险的累积效应迄今最大规模的精神分裂症GWAS(Trubetskoy et al., 2022,n>76,000例患者)已鉴定超过287个全基因组显著位点,涵盖基因包括DRD2(多巴胺D2受体)、CACNA1C(钙通道)、GRIN2A(NMDA受体亚基)等,反映多巴胺能与谷氨酸能系统的共同遗传基础。多基因风险评分(PRS)研究表明,PRS最高1%个体相比最低1%个体发病风险可增加40倍以上。
2.3 罕见变异:高效致病的遗传负担拷贝数变异(CNVs)
22q11.2缺失综合征携带者终生精神分裂症发病率约为25%–30%,OR超过60,是迄今已知最强的单一遗传风险因素。其他高风险CNVs包括1q21.1缺失、NRXN1缺失、16p11.2缺失等。CNV总体上解释约2%的精神分裂症遗传度,但在个体层面可产生决定性影响。
蛋白截短变异与有害错义变异
外显子组测序研究揭示,SETD1A(组蛋白甲基转移酶)、TRIO(Rho GEF,调控树突棘形态)、RB1CC1、SP4等基因的罕见功能丧失性变异显著增加精神分裂症风险。尤其值得关注的是,SETD1A的罕见截短变异OR约达12,提示表观遗传调控异常可能是精神分裂症发病的上游机制之一。
2.4 突触基因的功能汇聚尽管精神分裂症相关遗传变异在基因组层面分布广泛,但其编码产物在功能层面高度汇聚于突触信号传导网络,尤其是:
谷氨酸能突触:GRIN2A、GRIA3、GRM1等基因的变异提示NMDA受体信号通路的损伤,这与氯胺酮(NMDA拮抗剂)诱发精神病样症状的药理学证据高度一致
突触支架蛋白复合体:DLGAP2、MAGI2、SHANK3等基因维持突触后致密区的超分子结构,其功能缺失导致突触可塑性损伤
突触前囊泡释放:SNAP91、SV2A、DNM3等基因影响神经递质释放动力学
突触黏附分子:NRXN1-NLGN轴调控兴奋/抑制突触的形成与维持
2.5 遗传异质性与"遗传力缺失"精神分裂症遗传学研究面临的核心挑战是51%的遗传变异尚未归因于已知变异类型。可能的解释包括:基因-基因(上位性)互作、基因-环境互作、DNA甲基化等表观遗传变异、线粒体DNA变异、结构变异(倒位、易位)以及当前GWAS统计效力不足以捕获的亚阈值信号。未来研究需整合长读长测序、单细胞多组学及大规模跨种族队列以系统填补这一空白。